Agonistes ou antagonistes ? Telle est la question…
En se fixant sur leurs récepteurs, les molécules de communication intercellulaire de la famille des TGF-béta peuvent avoir des effets agonistes ou antagonistes, effets qui sont essentiels pour l’établissement des régions dorsales et ventrales de l’embryon. La base structurale de cette dichotomie entre agonistes et antagonistes est encore mal comprise. Dans un article publié dans PLOS Biology, des scientifiques révèlent que , dans le cas du TGF-beta d'oursin antagoniste PANDA, cette dichotomie repose sur la nature d’un seul acide aminé dans le domaine d’interaction du ligand à son récepteur.
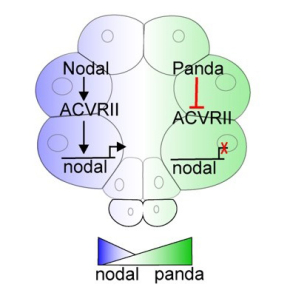
Panda: un TGF-beta au mécanisme d'action énigmatique
Chez l'oursin comme chez les mammifères, la restriction spatiale de l'expression du TGF-beta Nodal est l'élément clé qui brise la symétrie radiale de l'embryon et permet l'établissement de l'axe dorso-ventral. Cette restriction spatiale nécessite la production localisée de protéines de la famille des TGF-béta qui fonctionnent comme des antagonistes, se fixant sur le même récepteur que Nodal et bloquant sa voie de signalisation et son expression. Chez la souris, le produit du gène Lefty agit comme le premier facteur antagoniste de Nodal pour restreindre son expression. En revanche, chez l'oursin, c'est un facteur maternel (présent dès le stade œuf) nommé Panda (Paracentrotus Anti Nodal Dorsalizing Activity) qui agit comme antagoniste de Nodal afin de limiter son expression, Lefty n'intervenant que plus tardivement dans ce processus. Mais la base moléculaire de l'activité antagoniste de Panda n'était pas comprise.
La nature agoniste/antagoniste d'un TGF-beta peut-être codée au niveau d'un seul acide aminé
Dans un article publié dans Plos Biology les scientifiques lèvent en partie le voile sur le mécanisme par lequel le facteur Panda joue son rôle d'antagoniste de Nodal. Tout d'abord les scientifiques ont découvert que Panda inhibe fortement l'activité d'ACVRII, le récepteur sur lequel se fixe Nodal, en accord avec son rôle d'antagoniste. Ils se sont alors concentrés sur la région de Panda impliquée dans la liaison au récepteur ACVRII. En analysant les alignements de séquences de différents TGF-beta, ils ont fait une observation inattendue : dans les séquences de tous les facteurs Panda et Lefty de différentes espèces, c'est à dire chez quasiment tous les TGF-beta qui agissent comme des antagonistes, un acide aminé nommé proline est présent en position centrale dans la région impliquée dans l'interaction avec le récepteur ACVRII. En revanche, chez tous les autres TGF-beta agissant comme des agonistes (la majorité des BMPs, Nodal, Activin, GDF1-3 etc.), cet acide aminé est soit une sérine, soit une thréonine. Les scientifiques ont alors remplacé l'acide aminé proline par une sérine au sein de la séquence de Panda. Ce remplacement s'est avéré suffisant pour modifier radicalement l'activité de Panda sur Nodal qui est alors passée de celle d'un antagoniste favorisant l'acquisition d'une identité dorsale par les cellules en restreignant l'expression de nodal, à celle d'un agoniste de Nodal, promouvant l'acquisition d'une identité ventrale des cellules en stimulant l'expression de nodal.
L’acide aminé proline du domaine d’interaction entre Panda et ACVRII agirait comme un interrupteur moléculaire sur l’activité du récepteur.
Ces résultats montrent que le changement d'un acide aminé unique peut modifier de façon spectaculaire l'activité du TGF-beta PANDA. Ils suggèrent que la nature agoniste/antagoniste de PANDA, et probablement celle d'autres membres de la famille des TGF beta, est déterminée en grande partie par un seul acide aminé dans la région d'interaction du ligand avec le récepteur ACVRII. Cet acide aminé agirait comme un interrupteur moléculaire sur l'activité d’ACVRII : il activerait ACVRII lorsqu'il s'agirait d'une sérine (fonction agoniste), et l'inhiberait lorsqu'il s'agirait d'une proline (fonction antagoniste). Ces découvertes suggèrent qu'il pourrait être possible non seulement de prédire l'activité agoniste ou antagoniste d'un TGF-beta, mais aussi de convertir les antagonistes de la famille de Lefty en agonistes tels que Nodal, et les agonistes de la famille de Nodal/Activin en antagonistes tels que les Leftys, en modifiant la nature de l'acide aminé présent au sein du motif de liaison au récepteur ACVRII.
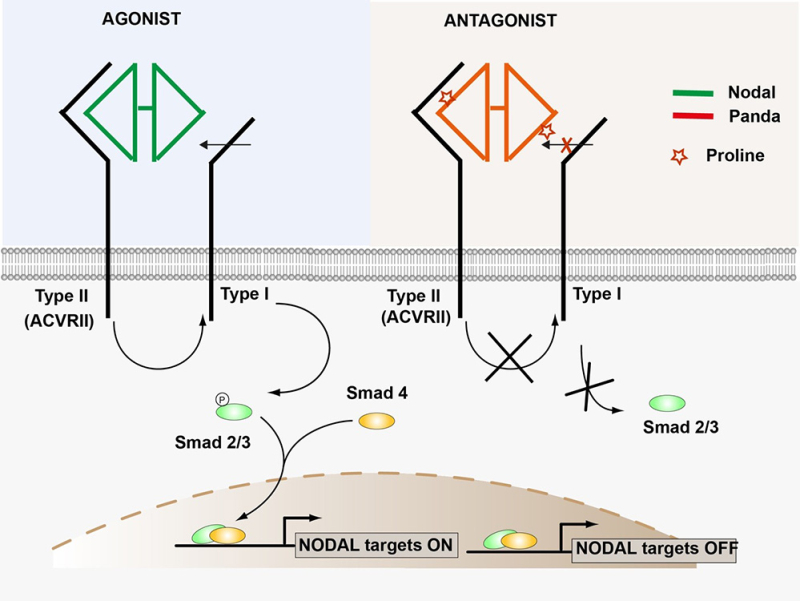
Figure : Le facteur Nodal se lie sous forme de dimère au récepteur ACVRII qui est constitutivement actif. Ceci permet le recrutement et la phosphorylation du récepteur de type I dans le complexe. Le récepteur de type I ainsi activé phosphoryle le facteur de transcription Smad2/3 ce qui active la transcription des gènes cibles de Nodal. La présence d'une Proline dans la région d'interaction entre Panda et ACVRII n'empêche pas la liaison entre Panda et le récepteur de type II mais elle bloque son activité et inhibe le recrutement du récepteur de type I ce qui inhibe l'activation de la transcription des gènes cibles.
Pour en savoir plus : Maternal TGF-β ligand Panda breaks the radial symmetry of the sea urchin embryo by antagonizing the Nodal type II receptor ACVRII
Viswanathan PK, Chessel A, Molina MD, Haillot E, Lepage T
PLOS Biology, 24 juin 2024, DOI : https://doi.org/10.1371/journal.pbio.3002701
Contact
Laboratoire
Institut de Biologie Valrose - IBV (CNRS/Inserm/Université Côte d'Azur)
UNIV COTE D'AZUR
Bâtiment Biochimie - Fac.de science
PARC VALROSE
06108 NICE