Des structures branchées !
Les introns de groupe II sont de grands ARN catalytiques et des éléments génétiques mobiles. Nés chez les bactéries, ils sont très probablement les ancêtres des introns nucléaires eucaryotes et de leur appareil d’épissage, le spliceosome. Dans les deux systèmes d’épissage, l’intron est excisé dans une conformation en ‘lasso’ due à une liaison spécifique 2’-5’. Des structures cristallographiques obtenues par des chercheurs de l’Institut de biologie intégrative de la cellule révèlent les raisons de l’extrême conservation de la forme ‘lasso’ au cours de l’évolution. Cette étude a été publiée le 2 décembre 2016 dans la revue Science.
© Maria Costa
© Maria Costa
En savoir plus
-
Crystal structures of a group II intron lariat primed for reverse splicing.
Costa M, Walbott H, Monachello D, Westhof E, Michel F.
Science. 2016 Dec 2;354(6316). pii: aaf9258.
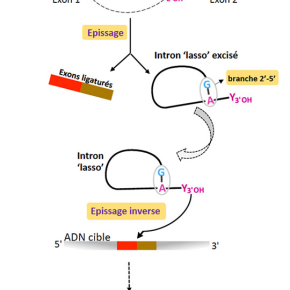
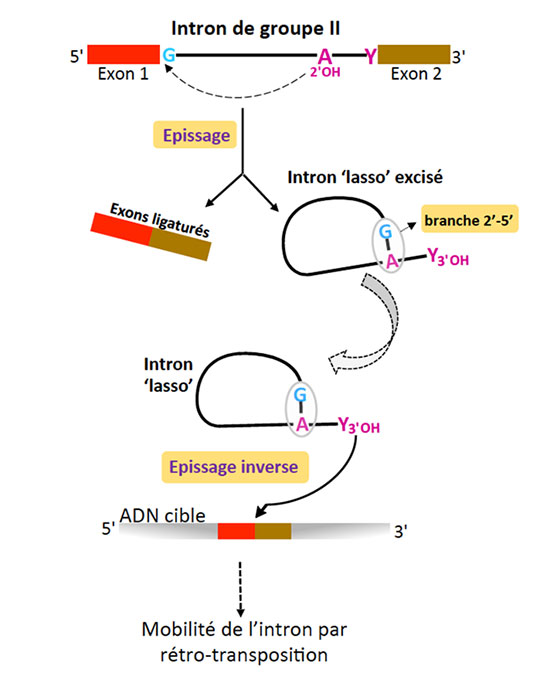
Les introns de groupe II sont de grandes molécules d’ARN capables de catalyser des réactions chimiques. Ce sont des ribozymes et cette propriété leur permet de s’auto-exciser d’un ARN précurseur (processus d’épissage) en l’absence de tout facteur protéique. Ces introns - très répandus chez les bactéries et dans les organites (mitochondrie et chloroplaste) - sont également des éléments génétiques mobiles, capables de coloniser des ADN cibles par un processus de rétro-transposition très efficacequi contribue à la diversification des génomes. Enfin, les introns de groupe II sont très probablement les ancêtres évolutifs des introns des ARN pré-messagers nucléaires eucaryotes et de leur machinerie d’épissage, le spliceosome. Cette hypothèse évolutive repose sur de nombreuses similarités au niveau structural et fonctionnel. Parmi celles-ci, la plus frappante est certainement le mécanisme d’épissage commun, qui aboutit à l’excision de l’intron sous forme d’une molécule branchée, appelée ‘lasso’ ou ‘lariat’. Dans les deux systèmes d’épissage, la conformation en ‘lasso’ résulte de la formation d’une liaison phosphodiester 2’-5’ entre une adénosine intronique très conservée et le premier nucléotide de l’intron, presque toujours une guanosine (figure 1).
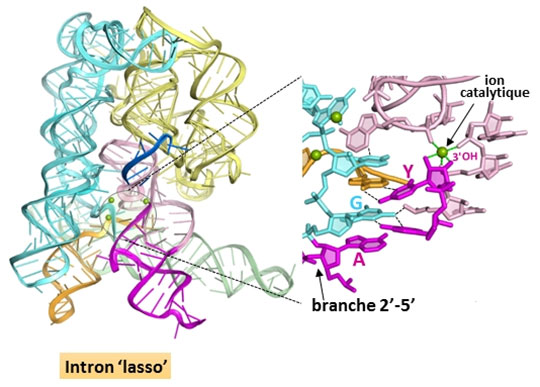
Trente ans après la découverte de la conformation ‘lasso’, on ne connaissait toujours pas les raisons de sa prédominance dans les systèmes d’épissage des organismes contemporains. Les structures cristallographiques d’un intron ‘lasso’ de groupe II obtenues par Maria Costa et ses collaborateurs illuminent cette question en révélant que la branche 2’(A)-5’(G) joue un rôle crucial dans l’assemblage du site actif de l’intron et dans la catalyse (figure 2). En mettant en interaction les nucléotides conservés aux deux extrémités de l’intron, la branche participe directement à la définition des jonctions d’épissage, accélère la réaction d’excision de l’intron par quelque trois ordres de grandeur et contribue de façon décisive à la fidélité du processus global d’épissage. De plus, ces structures montrent que le centre catalytique de l’intron lasso, une fois ce dernier libéré de l’ARN précurseur, est maintenu dans une configuration compétente pour l’initiation de « l’épissage inverse » (ou rétro-épissage) ; ce mécanisme, qui consiste en la catalyse des réactions d’épissage en sens inverse, est à la base de la propagation de l’intron dans les génomes par rétro-transposition. Ces données nouvelles fournissent une explication structurale à la supériorité de la forme ‘lasso’ en tant qu’élément mobile infectieux. Enfin, ces structures cimentent le lien évolutif entre les introns de groupe II et le système d’épissage des pré-messagers eucaryotes car elles suggèrent des homologies fonctionnelles précises entre nucléotides conservés dans les deux systèmes.
Des vecteurs dérivés des rétro-transposons de groupe II et dédiés à la manipulation du génome, sont déjà couramment utilisés chez les bactéries. Les connaissances tirées de ces travaux permettront la conception rationnelle de nouveaux vecteurs avec des propriétés modifiées ou inédites susceptibles, en autres, d’élargir le champ d’application de ces molécules à potentiel thérapeutique aux génomes eucaryotes.